This high-density forest biodiversity experiment (FAB1) focused on trees of our region investigates the consequences of multiple dimensions of tree diversity for soil processes, food webs, plant physiology, community dynamics, reproduction and mortality through time, and critical ecosystem functions, including productivity. FAB1 is specifically designed to unravel the ecological consequences of three forms of biological diversity: species richness (SR), functional variability (FV), and phylogenetic variability (PV). FV and PV are metrics of functional and phylogenetic diversity that are independent of species richness. We define FV as the representation of multiple traits of leaves, roots, seeds, and the whole organism responses to ennvironmental gradients of resource supply, growth, and decomposition. PV represents the variation among evolutionary lineages measured as the genetic distances between species. While PV and FV are often correlated, convergent evolution and adaptive differentiation can decouple them. When functional traits that drive specific ecosystem functions are not phylogenetically conserved, PV and FV may give contrasting predictions. PV, and FV are inherently not independent given shared ancestry among close relatives. We posit that PV and FV explain variation that SR cannot explain alone. Thus FAB is designed to examine the separate and combined effects of all three components of diversity for multiple ecosystem functions and to distinguish between “sampling” and “complementarity” effects of biodiversity. Due to the long lag between planting tree seedlings and determining effects of tree composition and diversity on ecosystem functioning, fewer experiments have been established to elucidate the role of biodiversity in the functioning of forest ecosystems than grassland experiments. FAB1 will contribute to address this gap and is a member of the IDENT and TreeDiv network of forest biodiversity experiments (www.treedivnet.ugent.be) and is critical the synthesis of long-term tree diversity experiments globally.
Hypotheses
Species and lineage effects
Productivity: Ecosystem function in monocultures will differ among plots planted with species or lineages that have contrasting functional trait values, particularly for “effect traits”, known to have ecosystem consequences. For example, we expect young monocultures to vary in productivity due to a range of growth strategies among species and lineages.
Decomposition and Nutrient Cycling: We also expect divergence in nutrient cycling due to contrasting properties of leaf and root detritus and exudates as well as mycorrhizal types. Due to shared ancestry that influences trait expression, effect traits will cause a phylogenetic signal in some ecosystem functions. Species and lineage effects on ecosystems resulting from chemical traits that influence soil processes are expected to become stronger through time. These effects will be stronger with increasing plot size due to reduced influence from neighboring assemblages.
Soil Community: Phylogenetically distinct monocultures will diverge in fungal and bacterial soil communities more than phylogenetically similar monocultures. This hypothesis is based on the postulate that shared ancestry results in similar decomposing root and leaf litter properties and similar fungal symbionts and soil pathogen communities as a consequence of host-specificity. These effects will become stronger through time.
Natural Enemies: Phylogenetically distinct monocultures will diverge in specialist herbivores, pathogens and enemy community compositions due to host specificity. These effects will become stronger as plot biomass increases through time and will increase with plot size due to greater host species apparency.
Mycorrhizal Type and Genotypic Effects: Fungal root and endophytic symbionts as well as specialist herbivores will show composition effects of host genotype within a species due to host specificity.
The FAB1 single species plots allow us to test hypotheses about the importance of plant functional traits in influencing ecosystem properties (e.g., NPP, soil C, N mineralization),plant-associated microbial communities, and individual tree ecophysiology. For example, we expect that plant species that increase concentrations of polyvalent soil cations (e.g., because of unique base cation chemistry or because of effects on soil acidity that influence Al and Fe solubility) will promote soil C stabilization through mineral-organic matter interactions and the formation of microaggregates that protect soil C from decomposition.

Experiment Design
from: Grossman, J. J., J. Cavender-Bares, S. E. Hobbie, P. B. Reich, and R. A. Montgomery. 2017. Species richness and traits predict overyielding in stem growth in an early-successional tree diversity experiment. Ecology 98:2601-2614. 10.1002/ecy.1958
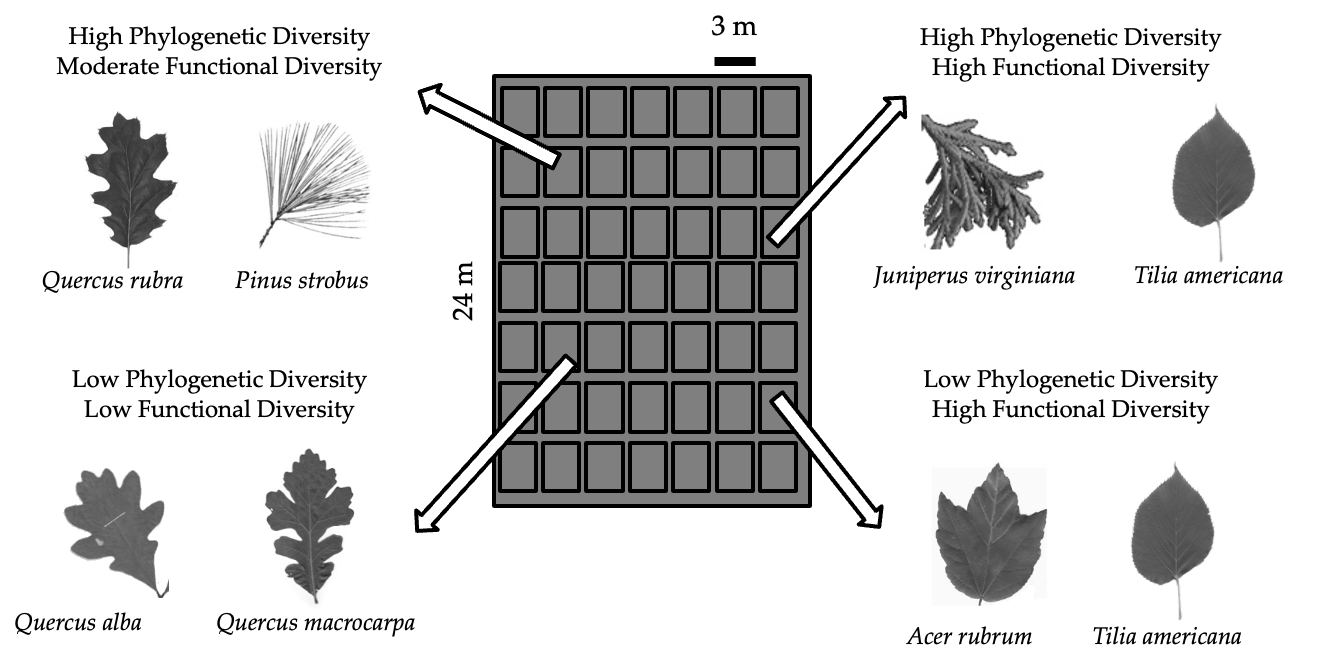
The FAB 1 High density diversity experiment consists of 8,960 trees of 12 native species. Four of these species are gymnosperms: eastern redcedar (Juniperus virginiana) and white (Pinus strobus), red (P. resinosa), and jack (P. banksiana) pine. The eight angiosperm species include red (Quercus rubra), pin (Q. ellipsoidalis), white (Q. alba), and bur (Q. macrocarpa) oak; red maple (Acer rubrum) and boxelder (A. negundo); paper birch (Betula papyrifera); and basswood (Tilia americana). Each of FAB`s three blocks (spaced 4.5 m apart) consists of either 46 or 47 square plots, each 3.5 m on the edge; plots are planted with one, two, five, or 12 species, with two-species plots additionally designed to tease apart functional and phylogenetic diversity. Each plot contains 64 trees, planted at 0.5 m intervals. Within a block, all trees are planted on a contiguous grid, without extra space in between plots. Each block contains 12 monocultural plots and 28 bicultural (two-species) plots; each of these plot types (or compositions) is therefore replicated three times across the experiment. Each block also contains either three or four random-draw five-species poly-cultures, the compositions of which are not replicated in the experiment, giving replication of the five-species level of richness but not of each five-species polyculture's composition. Each block also contains three or four 12-species polycultures, such that this composition is replicated 10 times across the experiment. Half of the 28 bicultural plot compositions were chosen by random draw. The other remaining bicultures were chosen using a stratified random approach designed to provide plots both low and high in PV and FV.
Planting and establishment
The experimental site was burned, then mulched with wood chips (from non-native western red cedar [Thuja plicata]) to prevent regrowth of herbaceous species. The experiment was planted over one week in late May 2013 with regionally sourced bare root seed-lings of unknown genetic relatedness that ranged from one to two years in age. Prior to planting, seedling roots were coated with commercial ectomycorrhizal and endomycorrhizal inoculum including species known to associate with all genera included in the experiment (Bio Organics, New Hope, PA). We used sprinkler irrigation to water newly planted seedlings ad libitum through June and July. We replanted seedlings as needed in May/June to maintain the plots; mortality was roughly 7 to 10 percent following replanting.
Treatment Tables
Download treatment tables (zip file)
Data
Select Publications
Species richness and traits predict overyielding in stem growth in an early‐successional tree diversity experiment
JJ Grossman, J Cavender‐Bares, SE Hobbie, PB Reich, RA Montgomery
2017 Ecology 98 (10), 2601-2614
Microbial community attributes supersede plant and soil parameters in predicting fungal necromass decomposition rates in a 12-tree species common garden experiment
F Maillard, B Beatty, M Park, S Adamczyk, B Adamczyk, CR See, ...
2023 Soil Biology and Biochemistry 184, 109124
Synthesis and future research directions linking tree diversity to growth, survival, and damage in a global network of tree diversity experiments
JJ Grossman, M Vanhellemont, N Barsoum, J Bauhus, H Bruelheide, ...
2018 Environmental and Experimental Botany 152, 68-89
Coupling spectral and resource-use complementarity in experimental grassland and forest communities
AK Schweiger, J Cavender-Bares, S Kothari, PA Townsend, MD Madritch, ...
2021 Proceedings of the Royal Society B 288 (1958), 20211290