Plant photosynthesis is the source of energy for most organisms on earth and plant biomass supports complex consumer foodwebs composed of a wide array of organisms including fungal pathogens, insect herbivores, and mammalian herbivores. In turn, these consumers can alter the productivity, diversity, and composition of the plant communities on which they depend. Many studies have the effects of examined individual groups of consumers through the use of herbivore fencing or insecticides, however few studies manipulate these groups in a single experiment. To quantify and compare the effects of multiple consumer groups, we established two experiments (E244 and E245) in 2008 that use insecticides, fungicides, and fences to manipulate the presence of insects, soil fungi, foliar fungi, or vertebrate herbivores. These experiments are conducted in two settings: an experiment that manipulates plant richness and composition and a natural, old field community. The first experiment (E244) is nested within 32 BigBio plots (E120) such that the fungicide and insecticide treatments are crossed with plant community diversity. The second experiment (E245) is conducted in an old field and has an additional treatment (unfenced, or fenced to exclude vertebrate herbivores). In 2019, we added a nutrient addition treatment to the old field plots (E245) at the subplot scale to quantify the interactions between nutrient supply and food web composition.
In both experiments, we apply treatments and measure NDVI weekly to bi-weekly throughout the growing season in all plots. We annually harvest biomass of each plant species, determine plant species diversity, and sample root biomass in all 253 experimental plots.
Experiment Design
from: Seabloom, E. W., Borer, E. T., & Kinkel, L. L. (2018; Sep) No evidence for trade-offs in plant responses to consumer food web manipulations. Ecology, 99(9), 1953-1963. doi:10.1002/ecy.2389
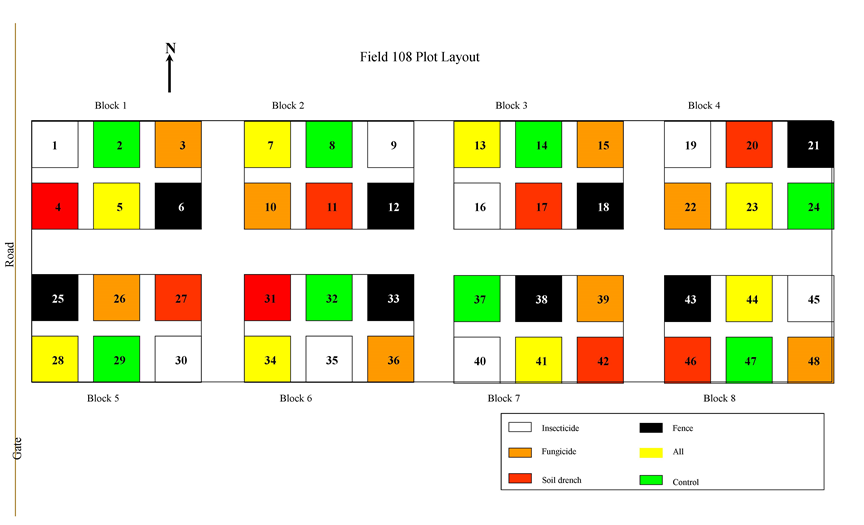
Plots layout
E245 was started in 2008 in an old-field abandoned from agriculture in 1956. The experiment is a completely randomized block design composed of 48 3 x 3 m plots arranged in eight blocks and randomly assigned to one of six treatments: control, insecticide, foliar fungicide, soil fungicide, fencing, or all treatments in combination (insecticide, foliar fungicide, soil fungicide, and fencing).
Fertilization
In 2019 a new treatment was imposed on the plots. In all the West subplots, fertilizer was added (nitrogen, phosphorus, potassium and micronutrients, all at a quantity of 100 grams per a meter squared).
Fencing
Plots in the fencing treatment are surrounded by 2 m tall wire fence with a 5 x 10 cm mesh to exclude moderate to large vertebrates; smaller vertebrates (e.g., pocket gophers and voles) had access to the plots.
Pesticide treatments
All pesticide treatments were applied throughout the duration of growing season, about mid-April to the end of August, each year. The foliar fungicide treatment is composed of twice monthly appli-cations of a combination of 7.5% Azoxystrobin and 2.5% Propiconazole (Quilt; Syngenta Crop Protection, Greens-boro, North Carolina, USA). The soil drench fungicide treatment is composed of monthly applications of 45.3%Mefenoxam (Ridomil Gold SL; Syngenta Crop Protection). The insecticide treatment was composed of twice monthly applications of 21.4% Imidacloprid (Marathon II; OHP, Mainland, Pennsylvania, USA). One to two times each growing season, Malathion is applied instead of Mara-thon II to limit the development of insecticide resistance in the insects.
Burning
The site is burned in the spring about every 3 years.
Treatment Tables
Download treatment tables (zip file)
Data
Select Publications:
No evidence for tradeoffs in plant responses to consumer food web manipulations
LK Seabloom, EW, ET Borer
2018 Ecology 99(9) 1953-1963
Stability of grassland production is robust to changes in the consumer food web
M Kohli, ET Borer, L Kinkel, EW Seabloom
2019 Ecology letters 22 (4), 707-716
Soil nutrients cause threefold increase in pathogen and herbivore impacts on grassland plant biomass
M Zaret, L Kinkel, ET Borer, EW Seabloom
2023 Journal of Ecology 111 (8), 1629-1640