Experiments 001 & 002 were established in 1982 to examine interactive effects of nitrogen addition (0-27.2 g N m-2 yr-1) and soil disturbance (tilling) on low-fertility grassland ecosystems. Both experiments have identical nutrient addition treatments, however E001 was started in an undisturbed grassland while E002 was disked with a tractor at the start of the experiment. This pair of experiments is replicated in three old fields (A, B, and C), and E001 is replicated in an additional field (D). These experiments were the first multi-decadal experiment to examine the impacts of chronic, experimental nitrogen addition as low as 10 kg of N per hectare per year above ambient atmospheric nitrogen deposition (6 kg of N per hectare per year). This total input rate is comparable to terrestrial nitrogen deposition in many industrialized nations.

Plots in E001 were sampled nearly annually for above-ground biomass (sorted by species) for the first two decades in all fields, however sampling is now periodic in fields A and B. The plots in E001 Field C comprise one of our two longest and most complete records of aboveground biomass and species composition, with the other being E054. Over the course of this experiment, new treatments have been imposed (Figure 1). In 1986, aluminum flashing was buried between the individual plots to prevent plants from spreading by vegetative reproduction. Starting in 2005, the successional grassland plots have been burned annually. The savanna plots have been burned every two out of three since 1987. See Figure 1. Timeline of treatments in E001. Gophers are trapped and removed from all experiment plots.
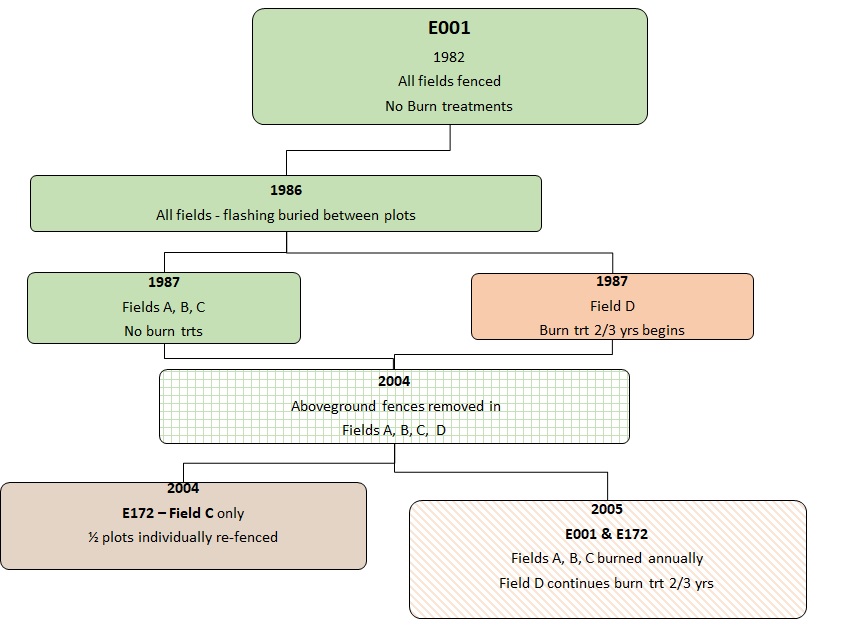
Figure 1. Timeline of treatments in E001
Experiment Design
from: From: Tilman, D. (1987), Secondary Succession and the Pattern of Plant Dominance Along Experimental Nitrogen Gradients. Ecological Monographs, 57: 189-214. https://doi.org/10.2307/2937080
Experiment E001 was established in four level, sandy, up-land, well-drained grassland fields within Cedar Creek Natural History Area, Minnesota. Three of the fields, A, B, and C, had been abandoned from farming 14, 25, and 48 yr, respectively, when this work began in 1982, and have never been burned. The fourth field, Field D, is a prairie opening within a stand of native oak savannah that has been maintained as oak savannah via prescribed bums in 2 out of every 3 yr, starting in 1966.
Nutrient Addition
Experimental nitrogen (N) gradients were established on existing vegetation in each field. A set of plots, each 4 x 4 m and separated by 1-m walkways, were established in Fields A, B, and C. The 54 plots per field were randomly assigned to receive one of nine different treatments, with six replicates per treatment per field. The treatments represent different combinations of nitrogen (0 – 27.2 g N m-2 yr-1 added as NH4NO3) and other non-N nutrients (20 g m-2 yr-1 P205; 20 g m-2 yr-1 K20; 40 g m-2 yr-1 CaCO3; 30.0 g m-2 yr-1 MgSO4; 18 mg m-2 yr-1 CuSO4; 37.7 mg m-2 yr-1 ZnSO4; 15.3 mg m-2 yr-1 CoCO2; 322 mg m-2 yr-1 MnCl2; and 15.1 mg m-2 yr-1 NaMoO4) applied annually. In this design, there is one treatment with no nutrients (Control; Treatment I) and eight treatments that received non-N nutrients in combination with eight different levels of N (0 – 27.2 g N m-2 yr-1). N applied at rates of 0, I, 2, 3.4, 5.4, 9.5, 17, or 27.2 g·m-2 -yr- 1 of N (Treatments A - H). Nitrogen is applied as commercial pelletized ammonium nitrate fertilizer (NPK content 34-0-0). Nutrients are added to plots in two equal doses each year, in early to mid-May and in late June, by manually broadcasting from the walk-ways. An identical experimental design was used in Field D, except that there were five replicates per treatment, and plots were 2 x 4 m. See Table 1. Tilman 1987
Fencing and flashing
The plots were originally enclosed by a fence to exclude mammalian herbivores. Galvanized welded-wire hardware cloth with 6mm x 6mm openings was buried to a depth of 84cm. Additional hardware cloth extended 60cm above the ground, and poultry netting extended to 1.8m above the ground. During the summer of 1986, aluminum flashing was placed in the ground to a depth of 12 inches. This flashing surrounds each plot in each field, keeping the plants from spreading between plots by below-ground vegetative reproduction.
Treatment Tables
Download treatment tables (zip file)
Data
Select Publications
Secondary succession and the pattern of plant dominance along experimental nitrogen gradients
D Tilman
1987 Ecological monographs 57 (3), 189-214
Biodiversity and stability in grasslands
D Tilman, JA Downing
1994 Nature 367 (6461), 363-365
Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands
CM Clark, D Tilman
2008 Nature 451 (7179), 712-715
Grassland ecosystem recovery after soil disturbance depends on nutrient supply rate
EW Seabloom, ET Borer, D Tilman
2020 Ecology Letters 23 (12), 1756-1765
Disturbance alters transience but nutrients determine equilibria during grassland succession with multiple global change drivers
MH DeSiervo, LL Sullivan, LM Kahan, EW Seabloom, LG Shoemaker
2023 Ecology letters